ЭНТРОПИЯ И ЭНЕРГИЯ В БИОЛОГИЧЕСКИХ СИСТЕМАХ. БИОФИЗИЧЕСКИЕ МЕХАНИЗМЫ АКТИВНОСТИ "ЭНЕРГЕТИЧЕСКИХ" МЕРИДИАНОВ
Коротков К. Г. 1, Виллиамс Б. 2, Виснески Л.А. 3
E-mail: korotkov@mail.admiral.ru
1 - СПбТУИТМО, Россия; 2 - Holos University Graduate Seminary, Fairview, Missouri; USA, 3-George Washington University Medical Center, USA.
Ведение
Методы исследования функционального состояния человека путем регистрации электро-оптических параметров кожного покрова можно разделить на две условные группы по характеру вовлекаемых биофизических процессов. К первой группе относятся "медленные" методы, время измерения в которых составляет более 1 с. При этом под влиянием приложенных потенциалов в тканях стимулируются ионно-деполяризационные токи и основной вклад в измеряемый сигнал вносит ионная компонента (Tiller, 1988). "Быстрые" методы, время измерения в которых составляет менее 100 мс, основаны на регистрации физических процессов, стимулированных электронной компонентой тканевой проводимости. Подобные процессы описываются в основном квантово-механическими моделями, поэтому их можно обозначить как методы квантовой биофизики. К последним относятся методы регистрации стимулированной и собственной люминесценции, а также метод стимулированной электронной эмиссии с усилением в газовом разряде (метод газоразрядной визуализации). Рассмотрим более детально биофизические и энтропийные механизмы реализации методов квантовой биофизики.
Электронная схема жизни
"Я глубоко убежден, что мы никогда не сможем понять сущность жизни, если ограничимся молекулярным уровнем… Удивительная тонкость биологических реакций обусловлена подвижностью электронов и объяснима только с позиций квантовой механики".
А. Сент-Дьердьи, 1971
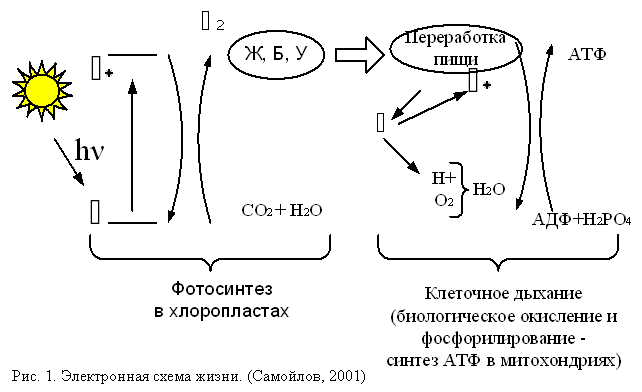
Электронная схема жизни - круговорот и превращение энергии в биологических системах, может быть представлена в следующем виде (Самойлов, 1986, 2001) (рис.1). Фотоны солнечного света поглощаются молекулами хлорофиллов, сосредоточенных в мембранах хлоропластов органоидов зеленых растений. Поглощая свет, электроны хлорофиллов приобретают дополнительную энергию и переходят из основного в возбужденное состояние. Благодаря упорядоченной организации белково-хлорофиллового комплекса, который носит название фотосистемы (ФС), возбужденный электрон не тратит энергию на тепловые превращения молекул, а приобретает способность преодолевать электростатическое отталкивание, хотя расположенное рядом с ним вещество имеет более высокий электронный потенциал, чем хлорофилл. В результате возбужденный электрон переходит на это вещество.
После потери своего электрона хлорофилл имеет свободную электронную вакансию. И он отбирает электрон у окружающих молекул, причем донором могут служить вещества, электроны которых имеют меньшую энергию, чем электроны хлорофилла. Этим веществом является вода (рис.2).
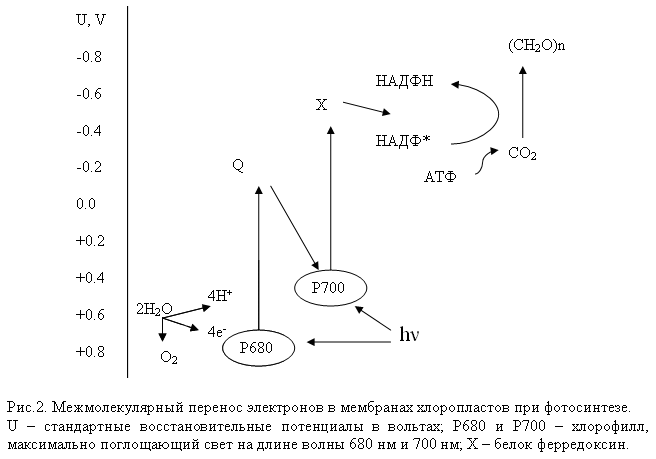
Отбирая электроны у воды, фотосистема окисляет ее до молекулярного кислорода. Так атмосфера Земли непрерывно обогащается кислородом.
При переносе подвижного электрона по цепи структурно связанных между собой макромолекул он тратит свою энергию на анаболические и катаболические процессы в растениях, а при соответствующих условиях, и у животных. По современным представлениям (Самойлов, 2001; Рубин, 1999) межмолекулярный перенос возбужденного электрона происходит по механизму туннельного эффекта в сильном электрическом поле.
Хлорофиллы служат промежуточной ступенькой в потенциальной яме между донором и акцептором электронов. Они принимают электроны от донора с низким энергетическим уровнем и за счет энергии солнца возбуждают их настолько, что они могут переходить на вещество с более высоким электронным потенциалом, чем донор. Это единственная, хотя и многоступенчатая световая реакция в процессе фотосинтеза. Дальнейшие аутотрофные биосинтетические реакции не нуждаются в свете. Они происходят в зеленых растениях за счет энергии, заключенной в электронах, принадлежащих НАДФН и АТФ. За счет колоссального притока электронов из двуокиси углерода, воды, нитратов, сульфатов и прочих сравнительно простых веществ создаются высокомолекулярные соединения: углеводы, белки, жиры, нуклеиновые кислоты.
Эти вещества служат основными питательными веществами для гетеротрофов. В ходе катаболических процессов, также обеспечиваемых электрон-транспортными системами, освобождаются электроны примерно в таком же количестве, в каком они захватывались органическими веществами при их фотосинтезе. Электроны, освобождаемые при катаболизме, переносятся на молекулярный кислород дыхательной цепью митохондрий (см. рис.1). Здесь окисление сопряжено с фосфорилированием - синтезом АТФ посредством присоединения к АДФ остатка фосфорной кислоты (то есть фосфорилирования АДФ). Этим обеспечивается энергоснабжение всех процессов жизнедеятельности животных и человека.
Находясь в клетке, биомолекулы "живут", обмениваясь энергией и зарядами, а значит, информацией, благодаря развитой системе делокализованных π-электронов. Делокализация означает, что единое облако π-электронов распределено определенным образом по всей структуре молекулярного комплекса. Это позволяет им мигрировать не только в пределах своей молекулы, но и переходить с молекулы на молекулу, если они структурно объединены в ансамбли. Явление межмолекулярного переноса было открыто J. Weiss в 1942 г., а квантовомеханическую модель этого процесса разработал в 1952-1964 гг R.S. Mulliken.
Вместе с тем важнейшая миссия π-электронов в биологических процессах связана не только с их делокализацией, но и с особенностями энергетического статуса: разность энергий основного и возбужденного состояний для них значительно меньше, чем у π-электронов и примерно равна энергии фотона hν.
Благодаря этому именно π-электроны способны аккумулировать и конвертировать солнечную энергию, за счет чего с ними связано все энергообеспечение биологических систем. Поэтому π-электроны принято называть "электронами жизни" (Самойлов, 2001).
Сопоставляя шкалы восстановительных потенциалов компонентов систем фотосинтеза и дыхательной цепи, нетрудно убедиться в том, что солнечная энергия, конвертированная π-электронами при фотосинтезе, затрачивается преимущественно на клеточное дыхание (синтез АТФ). Так, за счет поглощения двух фотонов в хлоропласте π-электроны переносятся от Р680 до ферредоксина (рис.2), увеличивая свою энергию примерно на 241 кДж/моль. Ее небольшая часть расходуется при переносе π-электронов с ферредоксина на НАДФ. В результате синтезируются вещества, которые затем становятся пищей для гетеротрофов и превращаются в субстраты клеточного дыхания. В начале дыхательной цепи запас свободной энергии π-электронов составляет 220 кДж/моль. Значит, до этого энергия π-электронов понизилась всего на 20 кДж/моль. Следовательно, более 90% солнечной энергии, запасенной π-электронами в зеленых растениях, доносится ими до дыхательной цепи митохондрий животных и человека.
Конечным продуктом окислительно-восстановительных реакций в дыхательной цепи митохондрий является вода. Она обладает наименьшей свободной энергией из всех биологически важных молекул. Говорят, будто с водой организм выделяет электроны, лишенные энергии в процессах жизнедеятельности. На самом деле запас энергии в воде отнюдь не нулевой, но вся энергия заключена в σ-связях и не может быть использована для химических превращений в организме при температуре тела и других физико-химических параметрах организма животных и человека. В этом смысле химическую активность воды принимают за точку отсчета (нулевой уровень) на шкале химической активности.
Из всех биологически важных веществ вода обладает самым высоким ионизационным потенциалом - 12,56 эВ. У всех молекул биосферы ионизационные потенциалы ниже этой величины, диапазон величин находится примерно в пределах 1 эВ (от 11,3 до 12,56 эВ).
Если принять ионизационный потенциал воды за точку отсчета реакционной способности биосферы, то можно построить шкалу биопотенциалов (рис.3). Биопотенциал каждого органического вещества имеет вполне определенное значение - он соответствует энергии, которая освобождается при окислении данного соединения до воды.

Размерность БП на рис.3 - это размерность свободной энергии соответствующих веществ (в ккал). И хотя 1 эВ = 1,6 10-19 Дж, при переходе от шкалы ионизационных потенциалов к шкале биопотенциалов нужно учитывать число Фарадея и разность стандартных восстановительных потенциалов между редокс-парой данного вещества и редокс-парой О2/H2O.
Благодаря поглощению фотонов электроны достигают наивысшего биопотенциала в фотосистемах растений. С этого высокого энергетического уровня они дискретно (по ступенькам) спускаются на самый низкий в биосфере энергетический уровень - уровень воды. Энергия, отдаваемая электронами на каждой ступеньке этой лестницы, превращается в энергию химических связей и таким образом движет жизнью животных и растений. Электроны воды связываются растениями, а клеточное дыхание вновь порождает воду. Этот процесс образует электронный кругооборот в биосфере, источником которого служит солнце.
Еще одним классом процессов, являющихся источником и резервуаром свободной энергии в организме, являются окислительные процессы, протекающие в организме с участием активных форм кислорода (АФК). АФК - это высоко реакционно-способные химические частицы, к которым относят содержащие кислород свободные радикалы (О2¾·, HО2·, НО·, NO·, ROO·), а также молекулы, способные легко продуцировать свободные радикалы (синглетный кислород, O3, ONOOH, HOCl, H2O2, ROOH, ROOR). В большинстве посвященных АФК публикаций, обсуждаются вопросы, связанные с их патогенным действием, поскольку долгое время считалось, что АФК появляются в организме при нарушениях нормального метаболизма, а в ходе инициированных свободными радикалами цепных реакций неспецифически повреждаются молекулярные компоненты клетки.
Однако сейчас стало ясно, что генерирующие супероксид ферменты имеются практически у всех клеток и что многие нормальные физиологические реакции клеток корре-лируют с увеличением продукции АФК [Sauer et al, 2001]. АФК генерируются и в ходе постоянно протекающих в организме неферментативных реакций [Mullarkey et al, 1990]. По минимальным оценкам, в состоянии покоя при дыхании человека и животных на производство АФК идет до 10-15% кислорода, а при повышении активности эта доля существенно возрастает [Лукьянова и др., 1982; Vlessis, et al., 1995]. При этом стационарный уровень АФК в органах и тканях в норме очень низок благодаря повсеместной распространенности мощных ферментативных и неферментативных устраняющих их систем. Вопрос о том, зачем организм столь интенсивно продуцирует АФК с тем, чтобы немедленно от них избавиться, в литературе пока не обсуждается.
Установлено, что адекватные реакции клетки на гормоны, нейромедиаторы, цитокины, на физические факторы (свет, температура, механические воздействия) требуют определенного содержании АФК в среде. АФК и сами могут вызывать в клетках те же реакции, что развиваются под действием биорегуляторных молекул - от активации или обратимого ингибирования ферментативных систем до регуляции активности генома. Биологическая активность так называемых аэроионов, оказывающих выраженное терапевтическое действие на широкий круг инфекционных и неинфекционных заболеваний [Чижевский, 1999], обусловлена тем, что они представляют собой свободные радикалы (О2¾·) [Goldstein, 1992]. Расширяется применение в терапевтических целях и других АФК - озона и перекиси водорода.
Важные результаты были получены в последние годы профессором Московского государственного университета В.Л. Воейковым [2003]. На основании большого объема экспериментальных данных по исследованию сверхслабого свечения цельной неразведенной крови человека было установлено, что в крови непрерывно протекают реакции с участием АФК, в ходе которых генерируются электронно-возбужденные состояния (ЭВС). Аналогичные процессы могут быть инициированы и модельных водных системах, содержащих аминокислоты и компоненты, способствующие медленному окислению аминокислот, в условиях близких к физиологическим. Энергия электронного возбуждения может излучательно и безызлучательно мигрировать в водных модельных системах и в крови, и использоваться как энергия активации для интенсификации процессов, порождающих ЭВС, в частности, за счет индукции вырожденного разветвления цепей.
Процессы с участием АФК, протекающие в крови и в водных системах, проявляют признаки самоорганизации, выражающиеся в их колебательном характере, устойчивости к действию интенсивных внешних факторов при сохранении высокой чувствительности к действию факторов низкой и сверх-низкой интенсивности. Это положение закладывает основу для объяснения многих эффектов, используемых в современной низкоинтенсивной терапии.
Полученные В.Л. Воейковым результаты демонстрируют еще один механизм генерации и утилизации ЭВС в организме, на этот раз в жидких средах. Развитие представлений, изложенных в данной главе, позволит обосновать биофизические механизмы генерации и транспорта энергии в биологических системах.
Энтропия жизни
В термодинамическом отношении открытые (биологические) системы в процессе функционирования проходят через ряд неравновесных состояний, что, в свою очередь, сопровождается изменением термодинамических переменных.
Поддержание неравновесных состояний в открытых системах возможно лишь за счет создания в них потоков вещества и энергии, что говорит о необходимости рассмотрения параметров таких систем как функции времени.
Изменение энтропии открытой системы может происходить за счет обмена с внешней средой (deS) и за счет роста энтропии в самой системе вследствие внутренних необратимых процессов (diS > 0). Э. Шредингер ввел понятие, что общее изменение энтропии открытой системы складывается из двух частей:
dS = deS + diS.
Продифференцировав это выражение, получим:
dS/dt = deS/dt + diS/dt.
Полученное выражение означает, что скорость изменения энтропии системы dS/dt равна скорости обмена энтропией между системой и окружающей средой плюс скорость возникновения энтропии внутри системы.
Член deS/dt , учитывающий процессы обмена энергией с окружающей средой, может быть и положительным, и отрицательным, так что при diS > 0 общая энтропия системы может как возрастать, так и убывать.
Отрицательная величина deS/dt < 0 соответствует тому, что отток положительной энтропии от системы во внешнюю среду превышает приток положительной энтропии извне, так что в результате общая величина баланса обмена энтропией между системой и средой является отрицательной. Очевидно, что скорость изменения общей энтропии системы может быть отрицательной при условии:
dS/dt < 0 if deS/dt < 0 and |deS/dt| > diS/dt.
Таким образом, энтропия открытой системы уменьшается за счет того, что в других участках внешней среды идут сопряженные процессы с образованием положительной энтропии.
Для земных организмов общий энергообмен можно упрощенно представить как образование в фотосинтезе сложных молекул углеводов из СО2 и Н2О с последующей деградацией продуктов фотосинтеза в процессах дыхания. Именно этот энергообмен обеспечивает существование и развитие как отдельных организмов - звеньев в круговороте энергии. Так и жизни на Земле в целом. С этой точки зрения уменьшение энтропии живых систем в процессе их жизнедеятельности обусловлено в конечном итоге поглощением квантов света фотосинтезирующими организмами, что, однако, с избытком компенсируется образованием положительной энтропии в недрах Солнца. Этот принцип относится и к отдельным организмам, для которых поступление извне питательных веществ, несущих приток "отрицательной" энтропии, всегда сопряжено с продуцированием положительной энтропии при их образовании в других участках внешней среды, так что суммарное изменение энтропии в системе организм + внешняя среда всегда положительно.
При неизменных внешних условиях в частично равновесной открытой системе в стационарном состоянии, близком к термодинамическому равновесию, скорость прироста энтропии за счет внутренних необратимых процессов достигает отличного от нуля постоянного минимального положительного значения.
diS/dt => Amin > 0
Этот принцип минимума прироста энтропии, или теорема Пригожина, представляет собой количественный критерий для определения общего направления самопроизвольных изменений в открытой системе вблизи равновесия.
Это условие можно представить по-другому:
d/dt (diS/dt) < 0
Это неравенство свидетельствует об устойчивости стационарного состояния. Действительно, если система находится в стационарном состоянии, то она не может самопроизвольно выйти из него за счет внутренних необратимых изменений. При отклонении от стационарного состояния в системе должны произойти внутренние процессы, возвращающие ее к стационарному состоянию, что соответствует принципу Ле-Шателье - устойчивости равновесных состояний. Иными словами, любое отклонение от устойчивого состояния вызовет увеличение скорости продуцирования энтропии.
В целом уменьшение энтропии живых систем происходит за счет свободной энергии, освобождаемой при распаде поглощаемых извне питательных веществ или за счет энергии солнца. Одновременно это приводит к увеличению их свободной энергии. Таким образом, поток отрицательной энтропии необходим для компенсации внутренних деструктивных процессов и убыли свободной энергии за счет самопроизвольных реакций метаболизма. В сущности, речь идет о круговороте и превращении свободной энергии, за счет которой поддерживается функционирование живых систем.
Диагностические технологии, основанные на достижениях квантовой биофизики
На основании рассмотренных выше представлений был развит целый ряд подходов, позволяющих исследовать прижизненную активность биологических систем. Это прежде всего спектральные методы, среди которых необходимо отметить методику одновременного измерения собственной флуоресценции НАДН и окисленных флавопротеидов (ФП), развитую коллективом авторов под руководством В.О. Самойлова. Эта методика основана на использовании оригинальной оптической схемы, разработанной Е.М. Брумбергом, позволяющей одномоментно измерять флуоресценцию НАДН на длине волны λ = 460 нм (синий свет) и флуоресценцию ФП на длине волны λ = 520-530 нм (желто-зеленый свет) при возбуждении ультрафиолетом (λ = 365 нм). В этой донорно-акцепторной паре донор π-электронов флуоресцирует в восстановленной форме (НАДН), а акцептор - в окисленной (ФП). Естественно, что в покое преобладают восстановленные формы, а при усилении окислительных процессов - окисленные.
Методика была доведена до практического уровня удобных эндоскопических приборов, что позволило разработать раннюю диагностику злокачественных заболеваний желудочно-кишечного тракта, лимфатических узлов в процессе хирургических операций, кожи. Принципиально важной оказалась оценка степени жизнеспособности тканей в процессе хирургических операций для проведения экономной резекции. Прижизненная флуометрия дает, кроме статических показателей, динамические характеристики биологических систем, так как позволяет проводить функциональные пробы и исследовать зависимость типа "доза-эффект". Это обеспечивает в клинике надежную функциональную диагностику и служит инструментом экспериментального изучения интимных механизмов патогенеза заболеваний.
К направлению квантовой биофизики можно отнести и метод газоразрядной визуализации (ГРВ). Стимулирование эмиссии электронов и фотонов с поверхности кожного покрова происходит за счет коротких (10 мкс) импульсов электромагнитного поля (ЭМП). Как показали измерения при помощи импульсного осциллографа с памятью, во время действия импульса ЭМП развивается серия импульсов тока (и свечения) длительностью примерно 10 нс каждый (рис.4). Развитие импульса обусловлено ионизацией молекул газовой среды за счет эмитированных электронов и фотонов, срыв импульса связан с процессами зарядки диэлектрической поверхности и возникновением градиента ЭМП, направленного противоположно исходному полю (Коротков, 2001). При подаче серии стимулирующих импульсов ЭМП с частотой следования 1000 Гц эмиссионные процессы развиваются в течение времени действия каждого импульса. Телевизионное наблюдение временной динамики свечения участка кожного покрова диаметром несколько миллиметров и покадровое сравнение картин свечения в каждом импульсе напряжения свидетельствует о возникновении эмиссионных центров практически в одних и тех же точках кожи.
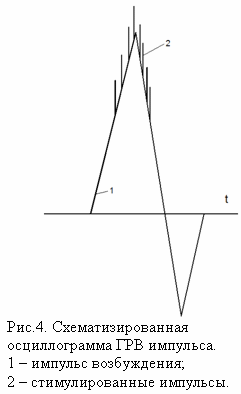 За столь короткое время - 10 нс - ионно-деполизационные процессы в ткани развиться не успевают, поэтому ток может быть обусловлен транспортом электронов по структурным комплексам кожи или иной исследуемой биологической ткани, включенной в цепь протекания импульсного электрического тока. Биологические ткани принято разделять на проводники (в первую очередь биологические проводящие жидкости) и диэлектрики. Для объяснения эффектов стимулированной электронной эмиссии необходимо рассматривать механизмы транспорта электронов по непроводящим структурам. Неоднократно высказывались идеи применить к биологическим тканям модель полупроводниковой проводимости. Полупроводниковая модель миграции электрона на большие межмолекулярные расстояния по зоне проводимости в кристаллической решетке хорошо известна и активно применяется в физике и технике. В соответствии с современными представлениями (Рубин, 1999), полупроводниковая концепция не получила подтверждения для биологических систем. В настоящее время наибольшее внимание в этой области привлекает к себе концепция туннельного транспорта электронов между отдельными белковыми молекулами-переносчиками, отделенными друг от друга энергетическими барьерами.
За столь короткое время - 10 нс - ионно-деполизационные процессы в ткани развиться не успевают, поэтому ток может быть обусловлен транспортом электронов по структурным комплексам кожи или иной исследуемой биологической ткани, включенной в цепь протекания импульсного электрического тока. Биологические ткани принято разделять на проводники (в первую очередь биологические проводящие жидкости) и диэлектрики. Для объяснения эффектов стимулированной электронной эмиссии необходимо рассматривать механизмы транспорта электронов по непроводящим структурам. Неоднократно высказывались идеи применить к биологическим тканям модель полупроводниковой проводимости. Полупроводниковая модель миграции электрона на большие межмолекулярные расстояния по зоне проводимости в кристаллической решетке хорошо известна и активно применяется в физике и технике. В соответствии с современными представлениями (Рубин, 1999), полупроводниковая концепция не получила подтверждения для биологических систем. В настоящее время наибольшее внимание в этой области привлекает к себе концепция туннельного транспорта электронов между отдельными белковыми молекулами-переносчиками, отделенными друг от друга энергетическими барьерами.
Процессы туннельного транспорта электронов хорошо экспериментально изучены и промоделированы на примере переноса электронов по белковой цепи. Туннельный механизм обеспечивает элементарный акт переноса электрона между донорно-акцепторными группами в белке, находящимися друг от друга на расстоянии порядка 0,5 - 1,0 нм. Однако существует много примеров, когда электрон переносится в белке на гораздо большие расстояния. Существенно, что при этом перенос происходит не только в пределах одной молекулы белка, но может включать взаимодействие разных белковых молекул. Так, в реакции переноса электрона между цитохромами с и цитохромом-оксидазой и цитохромом b5 оказалось, что расстояние между геммами взаимодействующих белков составляет более 2,5 нм (Рубин, 1999). Характерное время переноса электрона составляет 10-11 - 10-6 с, что соответствует времени развития единичного эмиссионного акта в методе ГРВ.
Проводимость белков может носить примесный характер. По данным экспериментов, значение подвижности u [м2/(В см)] в переменном электрическом поле составили для цитохрома ~ 1*10-4 , для гемоглобина ~ 2*10-4. В целом оказалось, что для большинства белков проводимость осуществляется в результате прыжков электронов между локализованными донорными и акцепторными состояниями, разделенными расстояниями в десятки нанометров. Лимитирующей стадией в процессе переноса является не движение заряда по токовым состояниям, а релаксационные процессы в доноре и акцепторе.
В последние годы удалось рассчитать реальные конфигурации такого рода "электронных троп" в конкретных белках. В этих моделях белковая среда между донором и акцептором разбивается на отдельные блоки, связанные между собой ковалентными и водородными связями, а также невалентными взаимодействиями на расстоянии порядка Ван-дер-вальсовых радиусов. Электронная тропа, таким образом, представляется комбинацией тех атомных электронных орбиталей, которые дают наибольший вклад в величину матричного элемента взаимодействия волновых функций компонентов.
В то же время общепризнанно, что конкретные пути переноса электрона не носят строго фиксированный характер. Они зависят от конформационного состояния белковой глобулы и могут соответственно меняться в различных условиях. В работах Маркуса был развит подход, в котором рассматривается не одна-единственная оптимальная траектория переноса в белке, а их набор. При вычислении константы переноса принимались во внимание орбитали целого ряда электронно-взаимодействующих атомов аминокислотных остатков белка между донорной и акцепторной группами, которые дают наибольший вклад в суперобменное взаимодействие. Оказалось, что для отдельных белков получаются более точные линейные зависимости, чем при учете одной-единственной траектории.
Трансформация электронной энергии в биоструктурах связана не только с переносом электронов, но и с миграцией энергии электронного возбуждения, которая не сопровождается отрывом электрона от молекулы донора. Наиболее важными для биологических систем, по современным представлениям, оказываются индуктивно-резонансный, обменно-резонансный и экситонный механизмы переноса электронного возбуждения. Эти процессы оказываются важными при рассмотрении процессов переноса энергии по молекулярным комплексам, как правило, не сопровождающихся переносом заряда.
Заключение
Рассмотренные представления показывают, что основным резервуаром свободной энергии в биологических системах являются электронно-возбужденные состояния сложных молекулярных комплексов. Эти состояния непрерывно поддерживаются за счет кругооборота электронов в биосфере, источником которого является солнечная энергия, а основным "рабочим веществом" - вода. Часть состояний тратится на обеспечение текущего энергоресурса организма, часть может запасаться впредь, подобно тому, как это происходит в лазерах после поглощения импульса накачки.
Протекание импульсного электрического тока в непроводящих биологических тканях может обеспечиваться за счет межмолекулярного переноса возбужденных электронов по механизму туннельного эффекта с активированным перескоком электронов в контактной области между макромолекулами. Таким образом, можно предположить, что формирование специфических структурно-белковых комплексов в толще эпидермиса и дермиса кожи обеспечивает формирование каналов повышенной электронной проводимости, экспериментально измеряемых на поверхности эпидермиса как электропунктурные точки. Гипотетически можно предположить наличие таких каналов и в толще соединительной ткани, что может быть ассоциировано с "энергетическими" меридианами. Иными словами, понятие переноса "энергии", характерное для представлений Восточной медицины и режущее слух человеку с европейским образованием, может быть ассоциировано с транс-портом электронно-возбужденных состояний по молекулярным белковым комплексам. При необходимости совершения физической или умственной работы в данной системе организма электроны, распределенные в белковых структурах, транспортируются в данное место и обеспечивают процесс окислительного фосфорилирования, то есть энергетического обеспечения функционирования локальной системы. Таким образом, организм формирует электронное "энергетическое депо", поддерживающее текущее функционирование и являющееся базисом для совершения работы, требующей мгновенной реализации огромных энергоресурсов или протекающей в условиях сверхбольших нагрузок, характерных, например, для профессионального спорта.
Стимулированная импульсная эмиссия также развивается в основном за счет транспорта делокализованных π-электронов, реализуемых в электрически непроводящей ткани путем туннельного механизма переноса электронов. Это позволяет предположить, что метод ГРВ позволяет косвенным образом судить об уровне энергетических запасов молекулярного уровня функционирования структурно-белковых комплексов.
Литература
|
Copyright © 1996-2022, Медицинская Академия Духовного Развития "МАДРА" При использовании представленной здесь информации ссылка на источник обязательна |
||
|
|
www.madra.dp.ua | Гостевая книга |
| Статистика посещаемости | Наш адрес | |
| видеоканал Dr. Evgen Semenikhin Become a Patron! | ||